
Обесценка "гормонов счастья"
#mental_health #neuroscience
Вдогонку к недавнему посту про "разоблачение" серотониновой теории депрессии следует подчеркнуть, что не только для ментальных заболеваний свойственны неоднородные проявления, которые невозможно описать дисфункцией одного нейромедиатора, но и роль самих нейромедиаторов многогранна. Поэтому описание серотонина или дофамина как гормонов счастья или удовольствия является не просто упрощением, но и искажением сути их функционирования.
Ниже представлены факты об этих двух нейромедиаторах, расширяющие традиционные представления о них.
Серотонин
- Несмотря на вовлеченность в функционирование нервной системы, 95% серотонина производится в кишечнике, где он вносит вклад в пищеварение.
- Серотонин участвует в выработке мелатонина — гормона, связанного с циркадными ритмами и циклами сна-бодрствования.
- Тромбоциты в крови содержат серотонин и направляют его к различным тканям, обеспечивая процессы заживления и свертывания крови.
- Регулируя обмен веществ, серотонин понижает аппетит и чувство голода, а также играет роль в поддержании тепла в организме.
- Большое количество серотонина может быть опасным: так называемый серотониновый синдром сопровождается повышением давления и частоты сердечных сокращений, тремором, проблемами с пищеварением. Осложнения могут приводить к судорогам, гиперрифлексии и даже смерти.
- Ваш мозг не может получить серотонин напрямую из еды. Сколько бы бананов вы ни съели, серотонин, содержащийся в них, не преодолеет гематоэнцефалический барьер и не попадет из кровяного русла в мозг. Но не стоит забывать про триптофан — аминокислоту, из которой образуется серотонин. Триптофан как раз может попасть из кровеносной системы в нервную, если вы будете употреблять еду с высоким содержанием белка.
Дофамин
- Дофамин не дарит вам эйфорию и удовольствие от той или иной деятельности. Он предсказывает ее исход. Если вам дают шоколадку, уровень выработки дофамина будет больше, если это преподношение окажется для вас неожиданным, в отличие от ситуации, когда вы уже ожидаете это вознаграждение. В обоих случаях при этом вы можете испытать одинаковое удовольствие от шоколада. Возможна и обратная ситуация: если вы ожидаете шоколадку, а вместо этого вас бьют током, содержание дофамина понизится. Это обеспечивает важные механизмы обучения: дофамин словно служит связующим звеном между вознаграждением и действиями, которые приводили к его получению. Он указывает вам на то, что вам понравилось, чтобы вы это повторили снова, а также на то, чего следует избегать. В этом —важное отличие между желанием и удовольствием.
- Дофамин регулирует выработку пролактина, отвечающего за репродуктивные функции: чем больше дофамина, тем меньше пролактина.
- Дофамин играет важную роль в регуляции движений. Разрушение дофаминергических нейронов сопровождает болезнь Паркинсона, нарушая баланс между активацией и торможением движения, что приводит к возникновению тремора.
- Избыток дофамина может вызывать негативные эффекты: тревожность, бессонницу, агрессию и даже галлюцинации.
#mental_health #neuroscience
Вдогонку к недавнему посту про "разоблачение" серотониновой теории депрессии следует подчеркнуть, что не только для ментальных заболеваний свойственны неоднородные проявления, которые невозможно описать дисфункцией одного нейромедиатора, но и роль самих нейромедиаторов многогранна. Поэтому описание серотонина или дофамина как гормонов счастья или удовольствия является не просто упрощением, но и искажением сути их функционирования.
Ниже представлены факты об этих двух нейромедиаторах, расширяющие традиционные представления о них.
Серотонин
- Несмотря на вовлеченность в функционирование нервной системы, 95% серотонина производится в кишечнике, где он вносит вклад в пищеварение.
- Серотонин участвует в выработке мелатонина — гормона, связанного с циркадными ритмами и циклами сна-бодрствования.
- Тромбоциты в крови содержат серотонин и направляют его к различным тканям, обеспечивая процессы заживления и свертывания крови.
- Регулируя обмен веществ, серотонин понижает аппетит и чувство голода, а также играет роль в поддержании тепла в организме.
- Большое количество серотонина может быть опасным: так называемый серотониновый синдром сопровождается повышением давления и частоты сердечных сокращений, тремором, проблемами с пищеварением. Осложнения могут приводить к судорогам, гиперрифлексии и даже смерти.
- Ваш мозг не может получить серотонин напрямую из еды. Сколько бы бананов вы ни съели, серотонин, содержащийся в них, не преодолеет гематоэнцефалический барьер и не попадет из кровяного русла в мозг. Но не стоит забывать про триптофан — аминокислоту, из которой образуется серотонин. Триптофан как раз может попасть из кровеносной системы в нервную, если вы будете употреблять еду с высоким содержанием белка.
Дофамин
- Дофамин не дарит вам эйфорию и удовольствие от той или иной деятельности. Он предсказывает ее исход. Если вам дают шоколадку, уровень выработки дофамина будет больше, если это преподношение окажется для вас неожиданным, в отличие от ситуации, когда вы уже ожидаете это вознаграждение. В обоих случаях при этом вы можете испытать одинаковое удовольствие от шоколада. Возможна и обратная ситуация: если вы ожидаете шоколадку, а вместо этого вас бьют током, содержание дофамина понизится. Это обеспечивает важные механизмы обучения: дофамин словно служит связующим звеном между вознаграждением и действиями, которые приводили к его получению. Он указывает вам на то, что вам понравилось, чтобы вы это повторили снова, а также на то, чего следует избегать. В этом —важное отличие между желанием и удовольствием.
- Дофамин регулирует выработку пролактина, отвечающего за репродуктивные функции: чем больше дофамина, тем меньше пролактина.
- Дофамин играет важную роль в регуляции движений. Разрушение дофаминергических нейронов сопровождает болезнь Паркинсона, нарушая баланс между активацией и торможением движения, что приводит к возникновению тремора.
- Избыток дофамина может вызывать негативные эффекты: тревожность, бессонницу, агрессию и даже галлюцинации.
{kind=link}
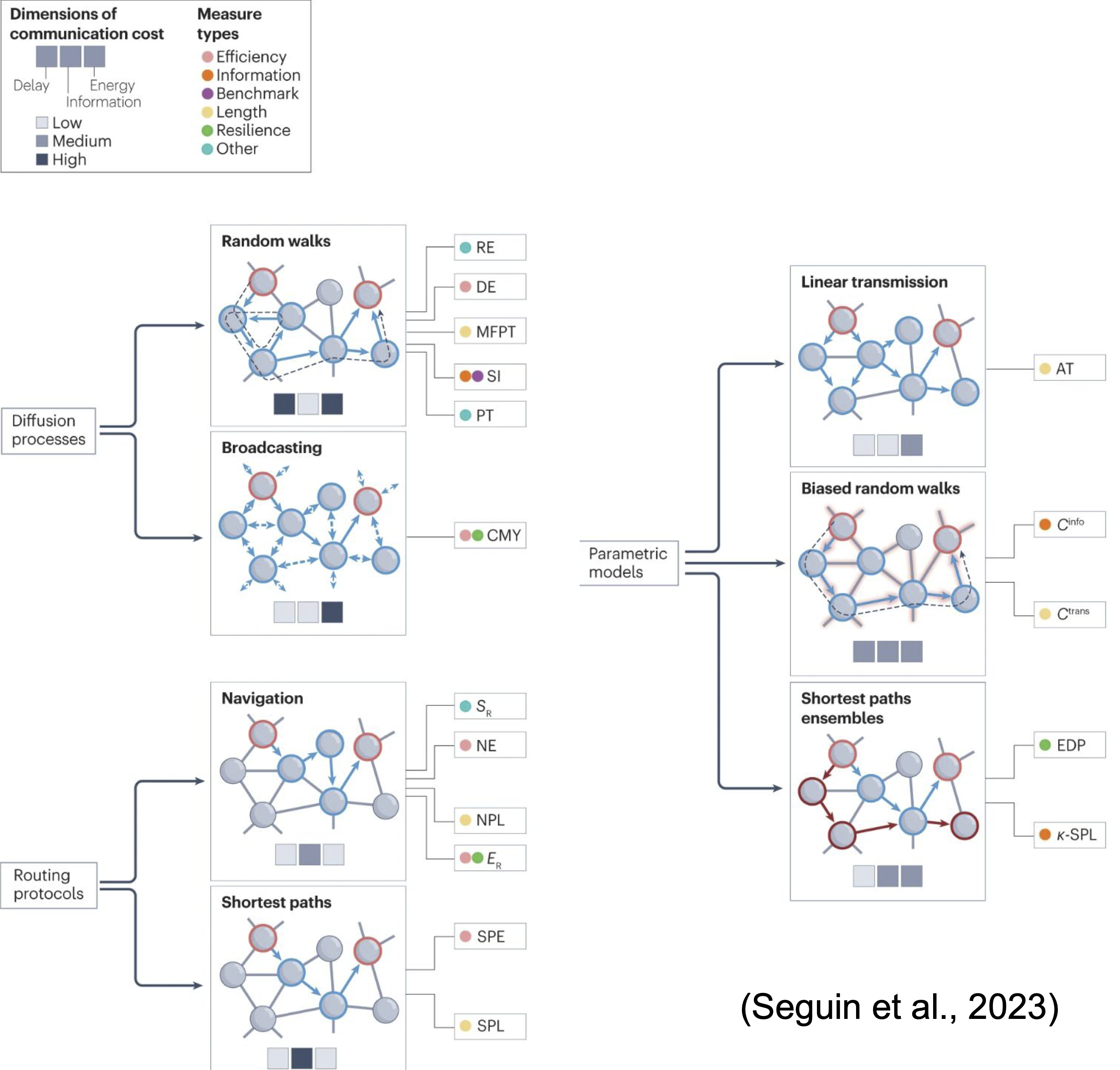
Такие разные коннектомы: новый обзор
#neuroscience
Публикация: Seguin, C., Sporns, O., & Zalesky, A. (2023). Brain network communication: concepts, models and applications. Nature Reviews Neuroscience, 1-18.
На днях был опубликован обзор, в котором систематизированы важные концепты коннектомики и сетевой нейронауки.
По своим свойствам коннектом – это small-world network, т. е. в этой сети пары далеких элементов разделены небольшим количеством промежуточных связей. Долгое время предполагалось, что нейронный сигнал распространяется по кратчайшим путям этой сети (shortest path assumption). По такой логике области мозга, анатомически связанные трактами белой материи, должны быть сильно связаны и функционально благодаря прямой передаче сигнала.
Но эксперименты показали, что это не совсем так. С одной стороны, структурная и функциональная связность действительно в среднем коррелируют. А с другой – области мозга, которые не связаны анатомически напрямую, тоже могут активно взаимодействовать. Более того, соблюдение ранее упомянутого критерия кратчайшего пути потребовало бы знания всей топологии сети. Но в случае с мозгом, сети которого децентрализованы, это не очень реалистичный сценарий: вряд ли индивидуальные нейроны или их группы обладают полной информацией о сети, в которой они находятся.
Раз принцип кратчайшего пути не срабатывает, то между структурным коннектомом (информацией об анатомических связях областей мозга) и функциональной связностью (информацией о том, насколько активность одной области мозга синхронизуется с другой) не хватает дополнительного звена. Этим звеном стали модели сетевой коммуникации – общие принципы распространения сигнала внутри структурного коннектома, которые и порождают те показатели функциональной связности, которые мы наблюдаем.
Семейства этих моделей, подробно описанные в обзоре, следующие:
1. Протоколы маршрутизации (routing protocols): нейрональные сигналы избирательно продвигаются по единственным надежным путям с низкой задержкой по времени и небольшими метаболическими затратами. Например, начиная с одного элемента сети, сигнал может перемещаться к соседнему, который наиболее близок к целевому элементу.
2. Процессы распространения: сигналы передаются диффузно, например, по принципу random walks или распространяясь сразу на несколько других элементов сети. Это увеличивает затрачиваемые ресурсы времени и энергии, но при этом позволяет ограничиваться информацией лишь о локальных связях. Кстати, в исследованиях по предсказанию функциональной связности на основе фМРТ с использованием структурного коннектома диффузные модели сработали лучше моделей кратчайшего пути.
3. Параметрические модели, реализующие гибридные стратегии предыдущих двух семейств.
Почему дальнейшая валидация этих групп моделей важна? Во-первых, их можно использовать для предсказания функциональных изменений вследствие нарушения структурного коннектома из-за травм, атрофии или хирургического вмешательства. Во-вторых, информацию о структурном коннектоме и о путях распространения сигнала в нем можно использовать для детального моделирования эффектов нейростимуляции, которая при фокальном воздействии на одну область может приводить к полисинаптическому ответу в анатомически не связанных областях.
Это не последний пост про структурные и функциональные связи в мозге и связь между ними (no pun intended): именно этой темой, пусть и несколько с другого ракурса, я занимаюсь в рамках своей диссертации. Обычно я не высказываюсь о ней научно-популярно из-за того, что, наверное, как и для многих аспирантов, диссертация становится чем-то многострадально-сакральным, о чем имеет смысл рассказывать либо все в мельчайших подробностях и перегружать читателя/слушателя, либо ничего. Но постараюсь найти баланс.
#neuroscience
Публикация: Seguin, C., Sporns, O., & Zalesky, A. (2023). Brain network communication: concepts, models and applications. Nature Reviews Neuroscience, 1-18.
На днях был опубликован обзор, в котором систематизированы важные концепты коннектомики и сетевой нейронауки.
По своим свойствам коннектом – это small-world network, т. е. в этой сети пары далеких элементов разделены небольшим количеством промежуточных связей. Долгое время предполагалось, что нейронный сигнал распространяется по кратчайшим путям этой сети (shortest path assumption). По такой логике области мозга, анатомически связанные трактами белой материи, должны быть сильно связаны и функционально благодаря прямой передаче сигнала.
Но эксперименты показали, что это не совсем так. С одной стороны, структурная и функциональная связность действительно в среднем коррелируют. А с другой – области мозга, которые не связаны анатомически напрямую, тоже могут активно взаимодействовать. Более того, соблюдение ранее упомянутого критерия кратчайшего пути потребовало бы знания всей топологии сети. Но в случае с мозгом, сети которого децентрализованы, это не очень реалистичный сценарий: вряд ли индивидуальные нейроны или их группы обладают полной информацией о сети, в которой они находятся.
Раз принцип кратчайшего пути не срабатывает, то между структурным коннектомом (информацией об анатомических связях областей мозга) и функциональной связностью (информацией о том, насколько активность одной области мозга синхронизуется с другой) не хватает дополнительного звена. Этим звеном стали модели сетевой коммуникации – общие принципы распространения сигнала внутри структурного коннектома, которые и порождают те показатели функциональной связности, которые мы наблюдаем.
Семейства этих моделей, подробно описанные в обзоре, следующие:
1. Протоколы маршрутизации (routing protocols): нейрональные сигналы избирательно продвигаются по единственным надежным путям с низкой задержкой по времени и небольшими метаболическими затратами. Например, начиная с одного элемента сети, сигнал может перемещаться к соседнему, который наиболее близок к целевому элементу.
2. Процессы распространения: сигналы передаются диффузно, например, по принципу random walks или распространяясь сразу на несколько других элементов сети. Это увеличивает затрачиваемые ресурсы времени и энергии, но при этом позволяет ограничиваться информацией лишь о локальных связях. Кстати, в исследованиях по предсказанию функциональной связности на основе фМРТ с использованием структурного коннектома диффузные модели сработали лучше моделей кратчайшего пути.
3. Параметрические модели, реализующие гибридные стратегии предыдущих двух семейств.
Почему дальнейшая валидация этих групп моделей важна? Во-первых, их можно использовать для предсказания функциональных изменений вследствие нарушения структурного коннектома из-за травм, атрофии или хирургического вмешательства. Во-вторых, информацию о структурном коннектоме и о путях распространения сигнала в нем можно использовать для детального моделирования эффектов нейростимуляции, которая при фокальном воздействии на одну область может приводить к полисинаптическому ответу в анатомически не связанных областях.
Это не последний пост про структурные и функциональные связи в мозге и связь между ними (no pun intended): именно этой темой, пусть и несколько с другого ракурса, я занимаюсь в рамках своей диссертации. Обычно я не высказываюсь о ней научно-популярно из-за того, что, наверное, как и для многих аспирантов, диссертация становится чем-то многострадально-сакральным, о чем имеет смысл рассказывать либо все в мельчайших подробностях и перегружать читателя/слушателя, либо ничего. Но постараюсь найти баланс.
{kind=link}
Мозг и его скорости
#neuroscience
Публикация: Senkowski, D., & Engel, A. K. (2024). Multi-timescale neural dynamics for multisensory integration. Nature Reviews Neuroscience, 1-18.
Наш мозг обрабатывает информацию, поступающую от разных органов чувств. Эта обработка не осуществляется изолированно: например, мы лучше понимаем речь (слуховая модальность), если наблюдаем за движением губ собеседника (зрительная модальность), и т. д. Объединение процессов такой обработки называется мультисенсорной интеграцией. Недавно в “Nature Reviews Neuroscience” появилась статья про её нейрональные механизмы.
Важной особенностью этого процесса является то, что он осуществляется на разных временных масштабах. Упрощённо: базовая сенсорная обработка происходит на небольших временных масштабах, а высокоуровневые процессы — на более длительных.
Что определяет эти временные масштабы? Синхронная активность нейронов в разных частотных диапазонах — ритмы мозга или осцилляции. Они задействуются в следующих сценариях.
✨ Модуляция амплитуды. Мультисенсорная интеграция сопровождается изменениями в мощности ритмов. Например, была установлена связь быстрой обработки мультисенсорных стимулов с мощностью гамма-ритма, а более сложных процессов (например, аудиовизуальной обработки речи) — с низкими частотами. Впрочем, это не означает, что иерархия мультисенсорной обработки линейно опирается на частоту таким образом, что высокие частоты связаны исключительно с простейшей сенсорной обработкой. Важно обращать внимание на то, какие области мозга генерируют эти ритмы: например, мультисенсорная интеграция может модулировать высокочастотные ритмы не только в сенсорных областях, но и в ассоциативных.
✨ Фазовый сброс (Phase resetting). Предъявление стимула в одной модальности может “обнулить” фазу осцилляций, отвечающих за процессы уже в другой модальности. Например, предъявление тактильного стимула обезьяне может приводить к сбросу фазы осцилляций в широком диапазоне частот (от 1 до 100 Гц) в слуховой коре, причем меньше чем через 100 мс после предъявления стимула. Этот механизм упреждающе может облегчать дальнейшую интеграцию предъявленного стимула в обработку уже другими системами.
✨ Ритмическая привязка (Entrainment). Стимуляция одной сенсорной системы на заданной частоте может индуцировать соответствующие осцилляции в другой системе. В частности, если предъявлять звуковые стимулы с частотой 3 Гц, можно наблюдать осцилляторную активность на этой же частоте в зрительной коре. А при предъявлении двух конкурирующих звуковых потоков целевые звуковые стимулы распознаются легче, если предъявлять зрительные стимулы, не относящиеся к задаче, но по временной динамике соотносимые с целевыми звуками.
✨ Функциональная связность. (Functional coupling). Осцилляции из функционально различных областей могут синхронизоваться на уровне фаз или огибающих. Например, синхронизация бета-ритма между височными и лобно-теменными областями наблюдается при эффекте Мак-Гурка (если вы не знакомы с этим эффектом, можете пройти по ссылке и мультисенсорно забыться). Важно также, что связность может устанавливаться и между разными ритмами: например, медленные осцилляции могут модулировать высокие.
Указанные выше механизмы касаются обработки уже предъявленного стимула. Но важно не забывать и про спонтанную активность незадолго до предъявления стимула. В частности, повышенная мощность спонтанного бета-ритма может предсказывать степень выраженности иллюзии Мак-Гурка, т. е. и предрасположенность к мультисенсорной интеграции в целом.
Также следует отметить, что в мозге имеют место процессы, не привязанные к временному масштабу (scale-free dynamics). Они не задействуют фиксированные частоты и не регулярны. Эту динамику связывают с состоянием критичности, в котором физическая система (мозг) наиболее подвержена изменениям во внешней среде. Соответственно, эти процессы могут влиять на то, с какой лёгкостью происходят ранее описанные фазовые сбросы или ритмические привязки, а также гибкость мультисенсорной интеграции в целом. Но это требует отдельных исследований.
#neuroscience
Публикация: Senkowski, D., & Engel, A. K. (2024). Multi-timescale neural dynamics for multisensory integration. Nature Reviews Neuroscience, 1-18.
Наш мозг обрабатывает информацию, поступающую от разных органов чувств. Эта обработка не осуществляется изолированно: например, мы лучше понимаем речь (слуховая модальность), если наблюдаем за движением губ собеседника (зрительная модальность), и т. д. Объединение процессов такой обработки называется мультисенсорной интеграцией. Недавно в “Nature Reviews Neuroscience” появилась статья про её нейрональные механизмы.
Важной особенностью этого процесса является то, что он осуществляется на разных временных масштабах. Упрощённо: базовая сенсорная обработка происходит на небольших временных масштабах, а высокоуровневые процессы — на более длительных.
Что определяет эти временные масштабы? Синхронная активность нейронов в разных частотных диапазонах — ритмы мозга или осцилляции. Они задействуются в следующих сценариях.
Указанные выше механизмы касаются обработки уже предъявленного стимула. Но важно не забывать и про спонтанную активность незадолго до предъявления стимула. В частности, повышенная мощность спонтанного бета-ритма может предсказывать степень выраженности иллюзии Мак-Гурка, т. е. и предрасположенность к мультисенсорной интеграции в целом.
Также следует отметить, что в мозге имеют место процессы, не привязанные к временному масштабу (scale-free dynamics). Они не задействуют фиксированные частоты и не регулярны. Эту динамику связывают с состоянием критичности, в котором физическая система (мозг) наиболее подвержена изменениям во внешней среде. Соответственно, эти процессы могут влиять на то, с какой лёгкостью происходят ранее описанные фазовые сбросы или ритмические привязки, а также гибкость мультисенсорной интеграции в целом. Но это требует отдельных исследований.
Please open Telegram to view this post
VIEW IN TELEGRAM